Michaelis-Menten Kinetics + some other stuff (highly relevant for USABO)
Michaelis-Menten kinetics describes the chemical kinetics of enzyme-catalyzed reactions in biological systems. Below is the equation:

The graph has substrate concentration, [S], on the x-axis, and reaction velocity on the y-axis. This equation assumes the simplest type of enzyme-substrate relation, i.e. E+S->ES complex, and ES complex->E+P (meaning that the enzyme and substrate get together, form a complex, and this complex returns the enzyme in its original form and a product).
The graph of Michaelis-Menten kinetics is below.

Notice that as substrate concentration increases, after a certain point, the velocity starts to level off. This is because in the reaction system, the amount of substrate present has exceeded the catalysis capacity of the enzymes that are present. This uncovers another key assumption of Michaelis-Menten kinetics: the concentration of enzyme is constant.
The velocity at which the graph levels off is the Vmax, or maximum reaction velocity given the constant amount of enzyme. The substrate concentration at which the velocity is half of the theoretical maximum velocity, Vmax, is called the Michaelis constant, or Km. Km is significant because it has an inverse relationship with enzyme affinity. This means that the lower the Km, the stronger the enzyme and substrate bind to one another, meaning that the enzyme works well at low substrate concentrations. This also makes sense with how the graph works, because if the Km is lower, it’s easier for the velocity to increase to that half point, and thus the enzyme is more effective. A high Km indicates that more substrate is required for the enzyme to function well (i.e. it’s harder for the enzyme to do a good job, if that rephrase makes sense).
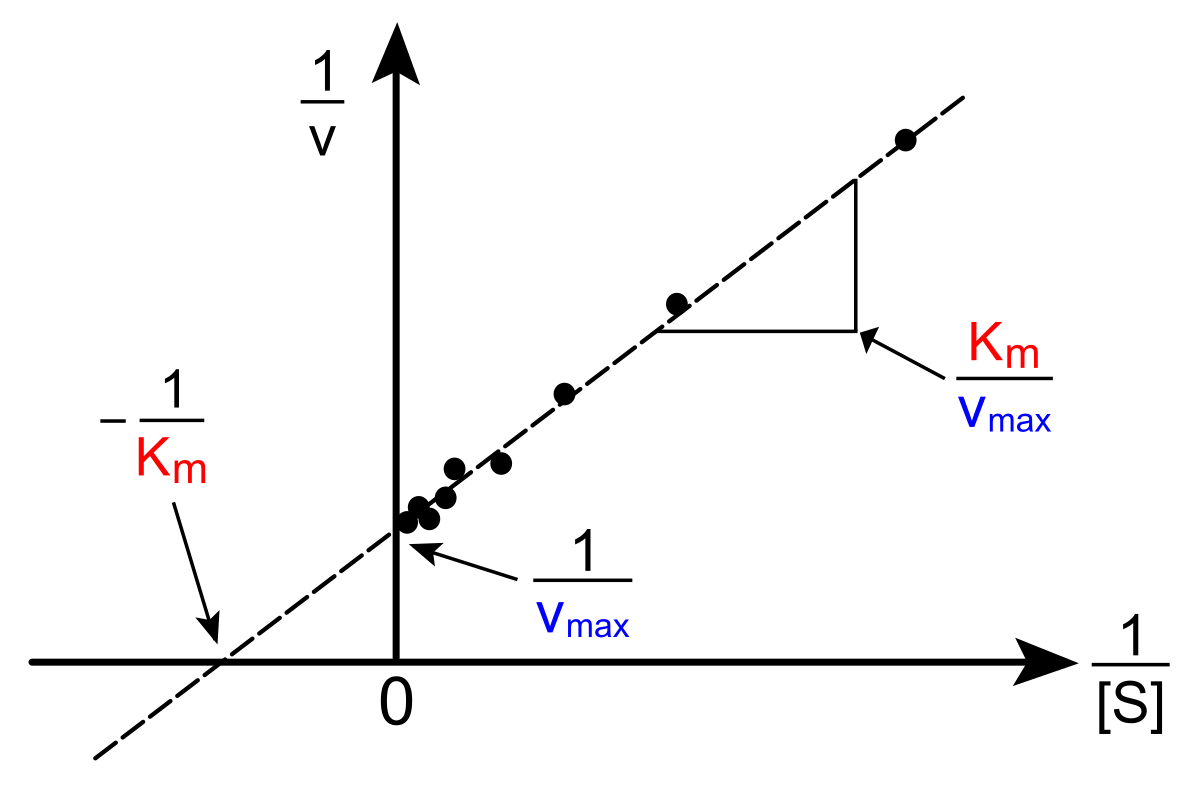
When we manipulate the Michaelis-Menten equation, we can create a linear version of this relationship, called the Lineweaver-Burk plot. The derivation is as follows: start with the original equation, and then:

take the reciprocal of both sides, split the sum, factor out the 1/[S], and there you have it! a linear equation in the form of y = mx + b, where y = 1/v, m = Km/Vmax, x = 1/[S], and b = 1/Vmax.
Here is what the Lineweaver-Burk plot looks like:
Remember how one assumption for these two equations is that the amount of enzyme is constant? Well, what if we encountered situations where some of the enzyme was rendered nonfunctional? This is exactly what happens with enzyme inhibition. There are three kinds of enzyme inhibition: competitive, noncompetitive, and uncompetitive.
Competitive inhibition occurs when the inhibitor binds to and blocks the enzyme’s active site meaning that the enzyme cannot bind to the substrate. This can be overcome by a large increase in substrate, so Vmax is not changed. However, because it slows down reaction progress, Km is increased (reflecting a lower enzyme-substrate affinity due to the blockage).
Noncompetitive inhibition is where the inhibitor binds to a site on the enzyme other than the active site to inactivate it (this site is the allosteric site). This decreases Vmax because the high substrate concentration cannot overcome this kind of inhibition. Km remains the same.
Finally, uncompetitive inhibition is where the inhibitor binds to the enzyme-substrate complex once it is formed, preventing product formation from the enzyme-substrate complex. Both Km and Vmax are decreased. Vmax is decreased because the maximal rate will be decreased and cannot be overcome by increased substrate. Km is decreased because the inability of the ES complex to be separated into product means that they are bound tightly, hence the affinity is increased.
Below are the Lineweaver-Burk plots for each of these scenarios, and descriptions of how each of the variables of the equation are modified:

So yeah that’s Michaelis-Menten and Lineweaver-Burk. Lmk if you have questions!